
南加州大学 Qi-Long Ying 团队 2026 年发表在《Cell》上的这项研究,直接把工程化髓系细胞的研发思路往前推了一大步 —— 他们盯上了造血分化路上的 “中间临时工”:粒细胞 - 单核细胞祖细胞(granulocyte-monocyte progenitors,GMPs),不仅用一套化学配方把它从 “传三代就分化” 变成了 “可长期规模化扩增” 的可再生平台,还设计出能跨越 MHC 屏障的 CAR-Fc 新结构,甚至在机制上挖出了一个颠覆认知的意外发现。
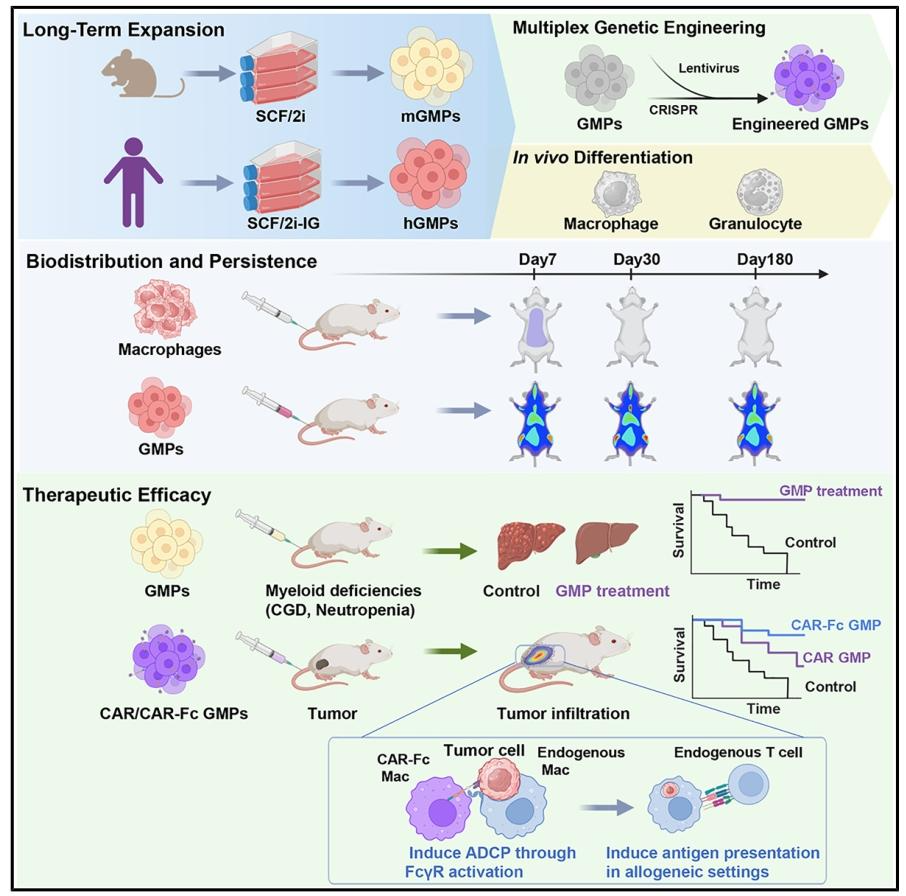
在经典造血发育理论里,GMP 是已经定向的髓系祖细胞,只能短暂扩增后就走向分化,从来没人把它当成可长期培养的工程化载体。
研究团队从小鼠体系入手,通过小分子筛选搭出了一套名为SCF/2i的无血清培养配方:干细胞因子 SCF + BRAF 抑制剂 GDC-0879 + 小分子 SKL2001。原代小鼠骨髓细胞丢进去,能连续稳定传代 50 天以上,细胞总量扩增近 8 个数量级,全程保持均一的未分化形态。更关键的是,扩出来的细胞 “身份没跑偏”:转录组、染色质开放区域、免疫表型都和原生 GMP 高度吻合,分化潜能也完整保留 —— 加 M-CSF 能变成有吞噬功能的巨噬细胞,加 G-CSF 能变成有杀菌能力的中性粒细胞,甚至还能定向分化为嗜碱性、嗜酸性粒细胞。
小鼠配方很完美,但放到人细胞上却 “水土不服”:SKL2001 对人源 GMP 的活性不足。团队直接对 SKL2001 做了构效优化,筛了 120 多个类似物,最终得到强效衍生物 TN2-30;再搭配 IL-3 和 G-CSF,升级出SCF/2i-IG人源专用配方。升级后的配方通吃所有常见起始材料:脐带血、成人骨髓、动员外周血来源的祖细胞都能稳定克隆扩增,连续培养 28 天后全基因组测序显示,没有突变富集、没有优势克隆爆发,基因组稳定性全程在线。
本文机制研究直接推翻了对一个经典小分子的认知。
SKL2001 在文献里一直被标注为 Wnt/β-catenin 通路激动剂,团队最初也默认它是靠激活 Wnt 来维持 GMP 增殖。但一系列实验结果直接推翻了这个假设:
1. 其他经典 Wnt 激动剂(CHIR99021、Wnt3a 等)全都复现不了 SKL2001 的扩增效果;
2. 加入 Wnt 通路抑制剂,也完全阻断不了 GMP 的增殖;
3. 最狠的证据:把 β-catenin 基因敲除的 GMP 放进 SCF/2i 培养基,细胞照样正常扩增。
结论很明确:SKL2001 维持 GMP 增殖,和经典 Wnt 通路一点关系都没有。
那它真正的靶点是什么?团队用带炔基标签的小分子衍生物做化学蛋白质组学钓靶,最终钓出来的核心靶点让所有人意外 ——髓过氧化物酶(MPO)。
MPO 是什么?是教科书里中性粒细胞的 “杀菌工具人”,靠催化产生次氯酸来消灭病原体,从来没人把它和髓系祖细胞的增殖调控联系在一起。为了实锤这个新功能,团队做了完整的验证链:
● GMP 中确实表达 MPO,且表达水平随分化动态变化;
● MPO 基因敲除的 GMP,不加 SKL2001 就比野生型有显著的增殖优势;一旦加入 SKL2001,这种优势就完全消失 —— 药物抑制和基因缺失的效果完全重合;
● 另外两种结构完全不同的 MPO 抑制剂(verdiperstat、mitiperstat),也能完美复现 GMP 长期扩增的效果;
● 表面等离子体共振(SPR)直接证明:SKL2001 可结合小鼠 MPO,TN2-30 可结合人 MPO,且二者都能直接抑制 MPO 的氯化酶活性。
这项研究顺手给 MPO 解锁了一个全新身份:髓系祖细胞增殖寿命与干性维持的关键调控因子。
CAR-M 临床前和早期临床的一大痛点,是静脉输注的巨噬细胞会被肺毛细血管截留,主要聚集在肺、肝、脾,肿瘤浸润量低,且一周左右就基本消失。GMP 作为上游祖细胞,完美解决了这个问题。
输注后,GMP 会主动定植到骨髓、脾脏等造血龛位,通过供体来源的髓系造血,源源不断产生单核细胞、巨噬细胞和粒细胞,再播散到全身组织,包括肿瘤微环境。定量数据对比非常直观:
● 直接输注的人源巨噬细胞,在小鼠体内不到 7 天就检测不到;
● 输注的人源 GMP 半衰期约 151 天,至少可维持 180 天;小鼠同源 GMP 半衰期约 142 天,持续存在超过 150 天。
功能也完全在线:在慢性肉芽肿病(CGD,吞噬细胞氧化杀菌功能缺陷的遗传病)小鼠模型中,输注扩增的 GMP 后,小鼠抵抗金黄色葡萄球菌、洋葱伯克霍尔德菌感染的能力大幅提升 —— 肝脓肿显著减少,脾脏大小恢复正常,生存率显著提高,相当于用体外扩增的祖细胞,临时重建了宿主的先天免疫防线。
成熟巨噬细胞增殖活性低,对病毒转导、CRISPR 编辑的兼容性很差,一直制约着 CAR-M 的工程化改造。而处于活跃增殖状态的 GMP,完美适配各类基因操作:慢病毒转导效率远高于成熟巨噬细胞,mRNA 转染、CRISPR-Cas9 基因敲除都能高效实现,甚至支持多轮序贯编辑,给多重工程化改造留足了空间。
团队分别构建了靶向 CD19 和 HER2 的 CAR-GMP,体外分化得到的 CAR 巨噬细胞,抗原特异性吞噬能力极强,联合 CD47 阻断抗体还能进一步提升吞噬效率。体内抗肿瘤效果同样亮眼:
● 血液瘤模型:CD19 CAR-GMP 输注后,60% 的白血病小鼠到第 80 天仍维持极低的肿瘤负荷,对照组小鼠全部在 38 天内达到终点;
● 实体瘤模型:HER2 CAR-GMP 几乎完全抑制了卵巢癌移植瘤的生长,生存期显著延长,肿瘤组织中出现大量供体来源的巨噬细胞浸润。
异体通用型细胞治疗有个核心瓶颈:MHC 不匹配导致供体细胞无法直接激活宿主 T 细胞,且自身会逐渐被免疫排斥。团队设计了一个极其巧妙的 CAR 结构升级:在胞外 scFv 和跨膜区之间,插入了一段 IgG1 的 Fc 结构域,命名为CAR-Fc。
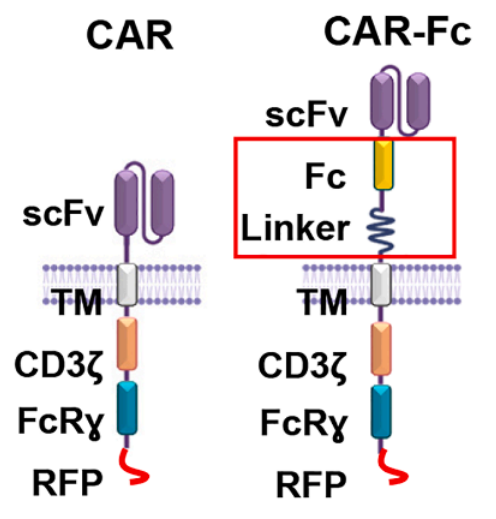
它的工作逻辑很精巧:CAR 结合肿瘤细胞表面抗原后,暴露的 Fc 段相当于给肿瘤细胞 “贴了一层抗体标签”。宿主自身表达 Fc 受体的吞噬细胞(比如树突状细胞)可以识别并吞噬这些靶细胞,再通过自身的 MHC 分子把肿瘤抗原递呈给 T 细胞 ——完全绕开了供体细胞的 MHC 限制,直接把抗原导入宿主 APC 的抗原呈递通路。
体外实验验证了这套机制:普通 CAR 只能介导供体细胞自身的 “顺式吞噬”,而 CAR-Fc 还能触发宿主吞噬细胞的 “反式吞噬”,并且能在完全 MHC 错配的情况下,激活抗原特异性 T 细胞的增殖和细胞因子分泌。到了体内异体肿瘤模型中,优势更加明显:
● 异体淋巴瘤模型:普通 CAR-GMP 仅能轻度延缓肿瘤进展,而 CAR-Fc 组大幅压制了肿瘤负荷,超过半数小鼠存活超过 60 天;
● 异体乳腺癌模型:CAR-Fc 组肿瘤内 CD3+T 细胞、颗粒酶 B + 效应 T 细胞的密度显著更高,肿瘤生长抑制更强,中位生存期较对照组接近翻倍。
更有意思的是,异体 GMP 本身会逐渐被宿主免疫系统清除,反而成了安全优势 —— 不会像 CAR-T 那样长期在体内存续,降低了脱靶毒性和长期组织损伤的风险,相当于 “精准起效、功成身退”。
这项工作最核心的颠覆,是打破了 “只有造血干细胞才能长期克隆扩增,定向祖细胞必然短命” 的固有认知 —— 只要找对了调控增殖与分化的内在刹车(比如 MPO),谱系定向的祖细胞也能成为稳定、可规模化的工程化平台。
从产业视角看,GMP 平台一次性解决了髓系细胞治疗 “扩增难、编辑难、体内存续差” 三大痛点;CAR-Fc 的设计又给异体免疫治疗提供了调动宿主免疫的全新思路。再加上 MPO 新功能的基础发现,无论对造血发育研究,还是对下一代细胞免疫治疗的开发,都是分量十足的突破。
Reference
 English
English


 18020283128
18020283128